This is the final paragraph in the discussion section of the paper by Skulachev, regarding the use of a Na+/K+ concentration gradient across a membrane to store potential energy, convertible to a Na+ or H+ gradient as needed, and why elevated K+ does not have to be a primordial feature of proto-cells:
"One might think that Na+ ions are incompatible with life and this is the reason why K+ is substituted for Na+ in the cell interior. Apparently, it is not the case as, e.g., in halophilic bacteria [Na+]int can reach 2 M [41]. The very fact that some enzyme systems work better in the presence of K+ than of Na+, may be considered as a secondary adaptation of enzymes to the K+-rich and Na+-poor conditions in the cytosol [40]. Besides, it would have been dangerous to couple any work performance with Na+ influx to the cytoplasm if Na+ were a cell poison".
That makes perfect sense to me.
Peter
Thursday, April 20, 2017
Wednesday, April 19, 2017
From Skulachev to LUCA
TLDR: Cells become islands of raised K+ ion concentration when energy is supplied.
Okay, here come the doodles based on Skulachev's paper
Membrane-linked energy buffering as the biological function of Na+/K+ gradient
This is the scenario in ultra modern bacteria, the pinnacle of about 4 billion years of evolution. The membrane is tight to all significant ions at reasonable temperatures and concentration gradients. In this set of pictures the proton population represented within the red circle is holding a membrane voltage of 180mV, as per usual:

The trans-membrane potential from the pumped protons is stable while ever the pumping and the consumption of protons is balanced. The problem is that it doesn't need many protons to generate that 180mV. Pumping any more than basic needs generates too great a membrane voltage. The converse is that it doesn't take much excess proton consumption to collapse the potential. So you need a buffer which does not waste the energy used to pump.
If a bacterium suddenly increases proton pumping by eating some glucose we have this problem of a spike in membrane voltage:

We can get around this by allowing a positive ion to travel in the opposite direction. This will stop the rising membrane potential as the ion uses the membrane potential to enter the cell against a concentration gradient. It uses an ion-specific channel, in this case for potassium. This process is electrophoresis down the electrical gradient, against a concentration gradient, powered by the electrical component rather than the pH component of the rising proton gradient:

The number of K+ ions matches the excess protons pumped. The electrical potential is thus maintained at 180mV at the "cost" or "benefit" (semantics here!) of K+ entering the cell. But there is a problem in that the more protons pumped and the more K+ entering the cell, the higher the pH of the intracellular medium becomes. That K+ pool is actually tied to the OH- left behind by pumping out H+. Caustic potash...

This is not good for metabolic processes. But it is easily surmounted using a 1:1 ratio Na+/H+ (electro-neutral) antiporter to get some protons back in to the cell to offset the excess OH-

while still maintaining an electrical gradient of 180mV using H+, keeping an electro-neutral Na+/K+ gradient as an energy store:

Obviously the Na+/H+ antiporter is being driven by the pH component of the proton gradient. It's neat how evolution has separated out the pH and electrical components of a proton gradient!
The whole system is fully reversible so if there is a sudden drop in proton pumping the transmembrane Na+/K+ gradient can be reconverted to a proton gradient to "buffer" changes in proton translocation. This seems to be how modern, proton pumping bacteria with superbly proton tight membranes work. In E coli the ion channel and antiporter are ATP gated.
That's how Skulachev looked at modern bacteria in 1978.
I'm now going to wander off on my own and speculate about LUCA with a proton leaky but Na+/K+ tight membrane. This is just me from here onwards:
Let's have a think about LUCA, with a cell membrane which is tight to Na+, and probably K+ too, but highly leaky to both protons and hydroxyl ions. Metabolism is based on Na+ pumping and a Na+ specific ATP synthase. The initial Na+/H+ antiporter (from the Life series) is gone as a source of Na+ gradient as soon as LUCA leaves the alkaline hydrothermal vents.
I like the idea that LUCA used a pyrophosphatase to pump Na+ but with any Na+ pump we have the same problem as in modern bacteria: You can only store a small amount of energy as a 180mV Na+ gradient, as per H+ above:

But excess Na+ pumping can be easily be accommodated by K+ electrophoresis:

There is no need for the Na+/H+ antiporter in this scenario because there is no pH change associated with pumping Na+ ions, so all we need is the ion specific channel for K+.
This sets up a non-electrical energy store which is "accessible" to form an electrical gradient when primary Na+ pumping is low.
The buffer automatically implies the generation of a raised intracellular K+. We have here, based on a tiny step beyond Skulachev's ideas, a place within LUCA which is potassium rich. It's simply produced to buffer changes in ion pumping by the primary Na+ pump (or usage by ATP synthase) across relatively primitive membranes. And driving intracellular K+ higher is an indicator to the cell that there is excess of energy available, which should select for increased enzyme activity based on rising intracellular K+ concentration. Many of the "core" LUCA enzymes do indeed use K+ as a cofactor to function optimally.
Summary: Cells become islands of raised K+ ion concentration when more than basal a level of energy is supplied. Remember that for our later discussion about Mulkidjanian's ideas on the origin of life on Earth.
Peter
Okay, here come the doodles based on Skulachev's paper
Membrane-linked energy buffering as the biological function of Na+/K+ gradient
This is the scenario in ultra modern bacteria, the pinnacle of about 4 billion years of evolution. The membrane is tight to all significant ions at reasonable temperatures and concentration gradients. In this set of pictures the proton population represented within the red circle is holding a membrane voltage of 180mV, as per usual:

The trans-membrane potential from the pumped protons is stable while ever the pumping and the consumption of protons is balanced. The problem is that it doesn't need many protons to generate that 180mV. Pumping any more than basic needs generates too great a membrane voltage. The converse is that it doesn't take much excess proton consumption to collapse the potential. So you need a buffer which does not waste the energy used to pump.
If a bacterium suddenly increases proton pumping by eating some glucose we have this problem of a spike in membrane voltage:

We can get around this by allowing a positive ion to travel in the opposite direction. This will stop the rising membrane potential as the ion uses the membrane potential to enter the cell against a concentration gradient. It uses an ion-specific channel, in this case for potassium. This process is electrophoresis down the electrical gradient, against a concentration gradient, powered by the electrical component rather than the pH component of the rising proton gradient:

The number of K+ ions matches the excess protons pumped. The electrical potential is thus maintained at 180mV at the "cost" or "benefit" (semantics here!) of K+ entering the cell. But there is a problem in that the more protons pumped and the more K+ entering the cell, the higher the pH of the intracellular medium becomes. That K+ pool is actually tied to the OH- left behind by pumping out H+. Caustic potash...

This is not good for metabolic processes. But it is easily surmounted using a 1:1 ratio Na+/H+ (electro-neutral) antiporter to get some protons back in to the cell to offset the excess OH-
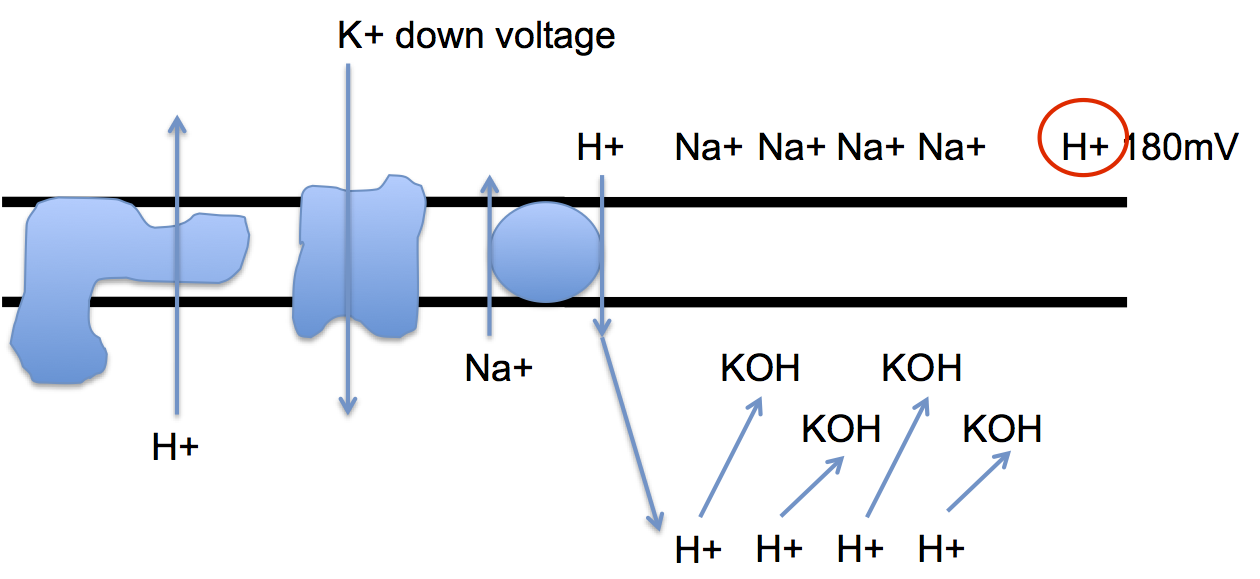
while still maintaining an electrical gradient of 180mV using H+, keeping an electro-neutral Na+/K+ gradient as an energy store:

Obviously the Na+/H+ antiporter is being driven by the pH component of the proton gradient. It's neat how evolution has separated out the pH and electrical components of a proton gradient!
The whole system is fully reversible so if there is a sudden drop in proton pumping the transmembrane Na+/K+ gradient can be reconverted to a proton gradient to "buffer" changes in proton translocation. This seems to be how modern, proton pumping bacteria with superbly proton tight membranes work. In E coli the ion channel and antiporter are ATP gated.
That's how Skulachev looked at modern bacteria in 1978.
I'm now going to wander off on my own and speculate about LUCA with a proton leaky but Na+/K+ tight membrane. This is just me from here onwards:
Let's have a think about LUCA, with a cell membrane which is tight to Na+, and probably K+ too, but highly leaky to both protons and hydroxyl ions. Metabolism is based on Na+ pumping and a Na+ specific ATP synthase. The initial Na+/H+ antiporter (from the Life series) is gone as a source of Na+ gradient as soon as LUCA leaves the alkaline hydrothermal vents.
I like the idea that LUCA used a pyrophosphatase to pump Na+ but with any Na+ pump we have the same problem as in modern bacteria: You can only store a small amount of energy as a 180mV Na+ gradient, as per H+ above:

But excess Na+ pumping can be easily be accommodated by K+ electrophoresis:

There is no need for the Na+/H+ antiporter in this scenario because there is no pH change associated with pumping Na+ ions, so all we need is the ion specific channel for K+.
This sets up a non-electrical energy store which is "accessible" to form an electrical gradient when primary Na+ pumping is low.
The buffer automatically implies the generation of a raised intracellular K+. We have here, based on a tiny step beyond Skulachev's ideas, a place within LUCA which is potassium rich. It's simply produced to buffer changes in ion pumping by the primary Na+ pump (or usage by ATP synthase) across relatively primitive membranes. And driving intracellular K+ higher is an indicator to the cell that there is excess of energy available, which should select for increased enzyme activity based on rising intracellular K+ concentration. Many of the "core" LUCA enzymes do indeed use K+ as a cofactor to function optimally.
Summary: Cells become islands of raised K+ ion concentration when more than basal a level of energy is supplied. Remember that for our later discussion about Mulkidjanian's ideas on the origin of life on Earth.
Peter
Monday, April 17, 2017
Skulachev in 1978
We know from papers like
Effect of Very Small Concentrations of Insulin on Forearm Metabolism. Persistence of Its Action on Potassium and Free Fatty Acids without Its Effect on Glucose
that, as we raise the concentration of insulin perfusing a tissue bed, the first effect is the suppression of lipolysis. Then it promotes potassium translocation in to cells. If you keep the concentration low enough there is zero effect on glucose translocation.
More practically: Anyone in first line general practice will be well familiar with the moribund cat with an obstructed bladder (thank you Go Cat) and a plasma K+ of 11.0mmol/l. You know the intravenous dose of Ca2+ you've given will stave off a-systole for a while and you've started to correct the acidosis with bicarbonate but the ECG still looks awful, as does the rest of the cat. Neutral insulin, covered by glucose, will usually drive potassium back in the cells where it belongs and keep the patient alive for long enough to allow you to get to work on the underlying problem. Pure potassium pragmatism.
So I have always wondered: Why does insulin facilitate active K+ translocation in to cells?
This strikes me as a very deep question. Always has.
There are hints as to why in Skulachev's paper from 1978.
Membrane-linked energy buffering as the biological function of Na+/K+ gradient.
I've only just found this paper and skimmed through it so far. It's a really interesting piece of theoretical bioenergetics from a close friend of the late Peter Mitchell. It was published in the year that Mitchell received his Nobel Prize for elucidating the principles of chemiosmosis. The paper is one of those which needs a note pad, a pencil and a pencil sharpener to work through. On the to-do list but I think it is saying that K+/Na+ translocation is an energy buffer to smooth out rapid changes in proton translocation energetics. That is a deep process.
I hope that's what Skulachev is saying!
And the follow on: Insulin signals a flood of calories. You're going to either spike delta psi or need to buffer it. That needs K+ to enter the cytoplasm to limit the voltage spike induced by the subsequent increase in H+ exit via pumping... Is insulin pre-empting this need? I'll try and get some doodles together but off-blog is getting busy at the moment.
Skulachev is still publishing important stuff today and his department is deeply involved in the evolutionary primacy of Na+ bioenergetics and, as a recent foray in to clinical pragmatism, the development of mitochondrial targeted antioxidants which appear to extend healthspan as well as lifespan.
Interesting chap and the 1978 paper strikes me as very perceptive and very prescient. You don't get many that good.
Peter
Effect of Very Small Concentrations of Insulin on Forearm Metabolism. Persistence of Its Action on Potassium and Free Fatty Acids without Its Effect on Glucose
that, as we raise the concentration of insulin perfusing a tissue bed, the first effect is the suppression of lipolysis. Then it promotes potassium translocation in to cells. If you keep the concentration low enough there is zero effect on glucose translocation.
More practically: Anyone in first line general practice will be well familiar with the moribund cat with an obstructed bladder (thank you Go Cat) and a plasma K+ of 11.0mmol/l. You know the intravenous dose of Ca2+ you've given will stave off a-systole for a while and you've started to correct the acidosis with bicarbonate but the ECG still looks awful, as does the rest of the cat. Neutral insulin, covered by glucose, will usually drive potassium back in the cells where it belongs and keep the patient alive for long enough to allow you to get to work on the underlying problem. Pure potassium pragmatism.
So I have always wondered: Why does insulin facilitate active K+ translocation in to cells?
This strikes me as a very deep question. Always has.
There are hints as to why in Skulachev's paper from 1978.
Membrane-linked energy buffering as the biological function of Na+/K+ gradient.
I've only just found this paper and skimmed through it so far. It's a really interesting piece of theoretical bioenergetics from a close friend of the late Peter Mitchell. It was published in the year that Mitchell received his Nobel Prize for elucidating the principles of chemiosmosis. The paper is one of those which needs a note pad, a pencil and a pencil sharpener to work through. On the to-do list but I think it is saying that K+/Na+ translocation is an energy buffer to smooth out rapid changes in proton translocation energetics. That is a deep process.
I hope that's what Skulachev is saying!
And the follow on: Insulin signals a flood of calories. You're going to either spike delta psi or need to buffer it. That needs K+ to enter the cytoplasm to limit the voltage spike induced by the subsequent increase in H+ exit via pumping... Is insulin pre-empting this need? I'll try and get some doodles together but off-blog is getting busy at the moment.
Skulachev is still publishing important stuff today and his department is deeply involved in the evolutionary primacy of Na+ bioenergetics and, as a recent foray in to clinical pragmatism, the development of mitochondrial targeted antioxidants which appear to extend healthspan as well as lifespan.
Interesting chap and the 1978 paper strikes me as very perceptive and very prescient. You don't get many that good.
Peter
Wednesday, April 05, 2017
Rho zero cells
Well, this post is about rho zero (ρ°) cells. TLDR: It's even more obscure than usual.
This is the basic ETC plus the ATP:ADP antiporter (ANT) and the Pi:H+ symporter (Slc25a3) added:

Most of this is very obvious but it's worth pointing out that ANT exchanges one ATP outwards with 4 negative charges for an ADP inwards which has 3 negative charges. The ADP needs an inorganic phosphate to reform ATP and this Pi carries one negative charge and enters the mitochondria via Slc25a3, facilitated by consuming one proton of the proton gradient. All is hunky dory with electrical balance, accepting some delta psi consumption.
ρ° cells are man made constructs which have no mitochondrial DNA, usually deleted by exposure to ethidium bromide. They live by glycolysis and need supplementary pyruvate and uridine to survive. They have no electron transport chain proteins because they lack core components needed to form complexes I, III, IV and the F0 (membrane) component of their F0F1 ATP synthase.
They do still form "petit" mitochondria. The F1 component of ATP synthase is present and it works. ANT and Slc25a3 are present and functional. There is CoQ, which is permanently reduced because there is nowhere for it to hand its electrons on to... A number of other cellular processes are also blocked, those which need to reduce CoQ to CoQH2 to occur. From
Restoration of electron transport without proton pumping in mammalian mitochondria
we have:

The really strange thing is that ρ° cells have a mitochondrial membrane potential and a proton gradient. This is what happens:

ATP which has been made in the cytoplasm enters the mitochondria via ANT running in reverse. The F1 component of the ATP synthase breaks down the ATP to ADP and Pi. ADP is exchanged outwards via the ANT antiporter and Pi is carried outwards in combination with a proton via the Slc25a3 symporter. This proton flux maintains the proton gradient across the inner mitochondria membrane, all of this process is being powered by glycolytic ATP synthesis.
I became interested in ρ° cells because the are so strange. But there are some practical things they tell us too. There's a venerable mini review here:
Cells depleted of mitochondrial DNA (ρ°) yield insight into physiological mechanisms
They cannot perform reverse electron transport through complex I, because there is no complex I. So no superoxide. Equally, there is none from complex III either. Clearly this has implications for what type of apoptosis they can perform and how they sense oxygen tension but more interestingly you can make ρ° versions of pancreatic beta cells.
These can't secrete insulin.
Back in the 1990s no one was thinking about RET as being essential to insulin secretion but they were pretty sure the process was based around mitochondria as well as needing glycolysis. In pancreatic beta cells glycolysis specifically inputs to the ETC at mtG3Pdh in large amounts, which will generate RET and the superoxide needed for insulin secretion. This occurs in other cells as part of insulin responsiveness, but not to the same degree as in the beta cells.
Placing some functional mitochondria in to ρ° beta cells restores insulin secretion ability.
The review suggests mtG3Pdh in beta cells acts as a sensor for cytoplasmic NADH levels. That's a nice idea. Just struck me as interesting.
Peter
This is the basic ETC plus the ATP:ADP antiporter (ANT) and the Pi:H+ symporter (Slc25a3) added:

Most of this is very obvious but it's worth pointing out that ANT exchanges one ATP outwards with 4 negative charges for an ADP inwards which has 3 negative charges. The ADP needs an inorganic phosphate to reform ATP and this Pi carries one negative charge and enters the mitochondria via Slc25a3, facilitated by consuming one proton of the proton gradient. All is hunky dory with electrical balance, accepting some delta psi consumption.
ρ° cells are man made constructs which have no mitochondrial DNA, usually deleted by exposure to ethidium bromide. They live by glycolysis and need supplementary pyruvate and uridine to survive. They have no electron transport chain proteins because they lack core components needed to form complexes I, III, IV and the F0 (membrane) component of their F0F1 ATP synthase.
They do still form "petit" mitochondria. The F1 component of ATP synthase is present and it works. ANT and Slc25a3 are present and functional. There is CoQ, which is permanently reduced because there is nowhere for it to hand its electrons on to... A number of other cellular processes are also blocked, those which need to reduce CoQ to CoQH2 to occur. From
Restoration of electron transport without proton pumping in mammalian mitochondria
we have:

The really strange thing is that ρ° cells have a mitochondrial membrane potential and a proton gradient. This is what happens:

ATP which has been made in the cytoplasm enters the mitochondria via ANT running in reverse. The F1 component of the ATP synthase breaks down the ATP to ADP and Pi. ADP is exchanged outwards via the ANT antiporter and Pi is carried outwards in combination with a proton via the Slc25a3 symporter. This proton flux maintains the proton gradient across the inner mitochondria membrane, all of this process is being powered by glycolytic ATP synthesis.
I became interested in ρ° cells because the are so strange. But there are some practical things they tell us too. There's a venerable mini review here:
Cells depleted of mitochondrial DNA (ρ°) yield insight into physiological mechanisms
They cannot perform reverse electron transport through complex I, because there is no complex I. So no superoxide. Equally, there is none from complex III either. Clearly this has implications for what type of apoptosis they can perform and how they sense oxygen tension but more interestingly you can make ρ° versions of pancreatic beta cells.
These can't secrete insulin.
Back in the 1990s no one was thinking about RET as being essential to insulin secretion but they were pretty sure the process was based around mitochondria as well as needing glycolysis. In pancreatic beta cells glycolysis specifically inputs to the ETC at mtG3Pdh in large amounts, which will generate RET and the superoxide needed for insulin secretion. This occurs in other cells as part of insulin responsiveness, but not to the same degree as in the beta cells.
Placing some functional mitochondria in to ρ° beta cells restores insulin secretion ability.
The review suggests mtG3Pdh in beta cells acts as a sensor for cytoplasmic NADH levels. That's a nice idea. Just struck me as interesting.
Peter
Saturday, April 01, 2017
Loki and its membrane potential
Nick Lane makes some interesting comments about Loki, currently accepted as being the closest living descendent of the archaeon which merged with an alpha proteobacterium to generate LECA, the Last Eukaryote Common Ancestor:
Lokiarchaeon is hydrogen dependent
Loki is fascinating. We don't quite have all of its genome, roughly 92% of it. There are bits missing for parts of ATP synthase and for the carbon monoxide dehydrogenase/acetyl-CoA synthase (CODH/ACS) complex but we can be pretty sure these are in that missing 8% of the genome which we have yet to find and sequence. After all, prokaryotes don't carry junk DNA. Having most of the genes for a functional complex suggests that the rest of those needed to make it work are present.
What is completely absent is anything suggesting any sort of respiratory chain. That's not so unusual, especially in anaerobes.
Or any sort of membrane pump.
No membrane pump? I suspect that there must be small ion pump of some sort either tucked away in the missing 8% of the genome or within some of the currently uninterpretable DNA. Certainly none of the large modern complex pumps are present in any part, so Ech, Rnf and MtrA-H are all out, although the MtrH gene alone is present. I'd assume MtrH is transferring methyl groups to somewhere other than to the absent MtrA-G Na+ pump, over to CODH/ACS seems most likely.
The core energy process appears to be based on using electron bifurcating hydrogenases to generate very low potential ferredoxins. This allows the CODH/ACS complex to generate acetyl phosphate or acetyl-CoA. Substrate level phosphorylation can then give ATP and it's all down hill, energetically speaking, from there onwards. This gives a strict anaerobic metabolism based on an external source of hydrogen.
Obviously a membrane gradient has many uses in addition to ATP synthesis so I wouldn't doubt for a moment that one is present. Keeping it energised is the trick.
It left me thinking about how you might generate a membrane potential in the absence of any obvious relative to modern day ion pumps. I recalled that Koonin had mentioned some very ancient sodium pumps based around either decarboxylation reactions or around pyrophosphate cleavage.
In Evolutionary primacy of sodium bioenergetics he comments:
"These ancestral ATPases [ATP synthase in reverse] would pump Na+ along with the Na+-transporting pyrophosphatase [62] and chemically-driven Na+-pumps, such as Na+-transporting decarboxylase [29,63], which, being found in both bacteria and archaea, appear to antedate the divergence of the three domains of life".
From which ref 62 is a good read
Na+-Pyrophosphatase: A Novel Primary Sodium Pump
"The role of Na+-PPase can be most easily conjectured in the thermophilic marine bacterium, T. maritima, which utilizes Na+ as the primary bioenergetic coupling ion and employs a Na+-ATP-synthase (35, 36). In this organism, Na+-PPase may work in concert with Na+-ATP-synthase to scavenge energy from biosynthetic waste (PPi) in order to maintain the Na+ gradient, especially under energy-limiting conditions".
And for Na+ pumping via conversion of succinate to proprionate:
Bacterial Na+- or H+-coupled ATP Synthases Operating at Low Electrochemical Potential
"A prominent example is Propionigenium modestum, which grows from the fermentation of succinate to propionate and CO2 (Schink and Pfennig, 1982; Dimroth and Schink, 1998). The free energy of this reaction is about -20 kJ/mol whereas approximately -70 kJ/mol is required to support ATP synthesis in growing bacteria (Thauer et al., 1977). To solve this apparent paradox, 3–4 succinate molecules must be converted into propionate before one ATP molecule can be synthesized".
This last process is somewhat more complex than pyrophosphate hydrolysis and looks less of a candidate for "hidden" membrane potential generation than the Na+PPase. After all, CODH/ACS is providing ATP and many reactions which need to be "one-way" cleave ATP to AMP and PPi. The PPi "waste" would then be available to pump Na+.
My guess would be that Loki will turn out to use Na+ membrane energetics...
Time will tell.
Peter
Lokiarchaeon is hydrogen dependent
Loki is fascinating. We don't quite have all of its genome, roughly 92% of it. There are bits missing for parts of ATP synthase and for the carbon monoxide dehydrogenase/acetyl-CoA synthase (CODH/ACS) complex but we can be pretty sure these are in that missing 8% of the genome which we have yet to find and sequence. After all, prokaryotes don't carry junk DNA. Having most of the genes for a functional complex suggests that the rest of those needed to make it work are present.
What is completely absent is anything suggesting any sort of respiratory chain. That's not so unusual, especially in anaerobes.
Or any sort of membrane pump.
No membrane pump? I suspect that there must be small ion pump of some sort either tucked away in the missing 8% of the genome or within some of the currently uninterpretable DNA. Certainly none of the large modern complex pumps are present in any part, so Ech, Rnf and MtrA-H are all out, although the MtrH gene alone is present. I'd assume MtrH is transferring methyl groups to somewhere other than to the absent MtrA-G Na+ pump, over to CODH/ACS seems most likely.
The core energy process appears to be based on using electron bifurcating hydrogenases to generate very low potential ferredoxins. This allows the CODH/ACS complex to generate acetyl phosphate or acetyl-CoA. Substrate level phosphorylation can then give ATP and it's all down hill, energetically speaking, from there onwards. This gives a strict anaerobic metabolism based on an external source of hydrogen.
Obviously a membrane gradient has many uses in addition to ATP synthesis so I wouldn't doubt for a moment that one is present. Keeping it energised is the trick.
It left me thinking about how you might generate a membrane potential in the absence of any obvious relative to modern day ion pumps. I recalled that Koonin had mentioned some very ancient sodium pumps based around either decarboxylation reactions or around pyrophosphate cleavage.
In Evolutionary primacy of sodium bioenergetics he comments:
"These ancestral ATPases [ATP synthase in reverse] would pump Na+ along with the Na+-transporting pyrophosphatase [62] and chemically-driven Na+-pumps, such as Na+-transporting decarboxylase [29,63], which, being found in both bacteria and archaea, appear to antedate the divergence of the three domains of life".
From which ref 62 is a good read
Na+-Pyrophosphatase: A Novel Primary Sodium Pump
"The role of Na+-PPase can be most easily conjectured in the thermophilic marine bacterium, T. maritima, which utilizes Na+ as the primary bioenergetic coupling ion and employs a Na+-ATP-synthase (35, 36). In this organism, Na+-PPase may work in concert with Na+-ATP-synthase to scavenge energy from biosynthetic waste (PPi) in order to maintain the Na+ gradient, especially under energy-limiting conditions".
And for Na+ pumping via conversion of succinate to proprionate:
Bacterial Na+- or H+-coupled ATP Synthases Operating at Low Electrochemical Potential
"A prominent example is Propionigenium modestum, which grows from the fermentation of succinate to propionate and CO2 (Schink and Pfennig, 1982; Dimroth and Schink, 1998). The free energy of this reaction is about -20 kJ/mol whereas approximately -70 kJ/mol is required to support ATP synthesis in growing bacteria (Thauer et al., 1977). To solve this apparent paradox, 3–4 succinate molecules must be converted into propionate before one ATP molecule can be synthesized".
This last process is somewhat more complex than pyrophosphate hydrolysis and looks less of a candidate for "hidden" membrane potential generation than the Na+PPase. After all, CODH/ACS is providing ATP and many reactions which need to be "one-way" cleave ATP to AMP and PPi. The PPi "waste" would then be available to pump Na+.
My guess would be that Loki will turn out to use Na+ membrane energetics...
Time will tell.
Peter
Subscribe to:
Posts (Atom)
